
Larval Import into the Hawaiian Archipelago
Many marine organisms exhibit biphasic life histories with a small, pelagic early life stage. Organisms with a second benthic life stage rely on larval dispersal for connectivity among populations. Even for many organisms with a second mobile life stage, the larval stage is often still the dominant stage for dispersal. Larval dispersal potential, and therefore connectivity, is controlled by a variety of physical (i.e. currents, dispersion, temperature) and biological (i.e. PLD, timing of spawning, larval behavior, mortality) factors. These factors vary greatly over both time and space.
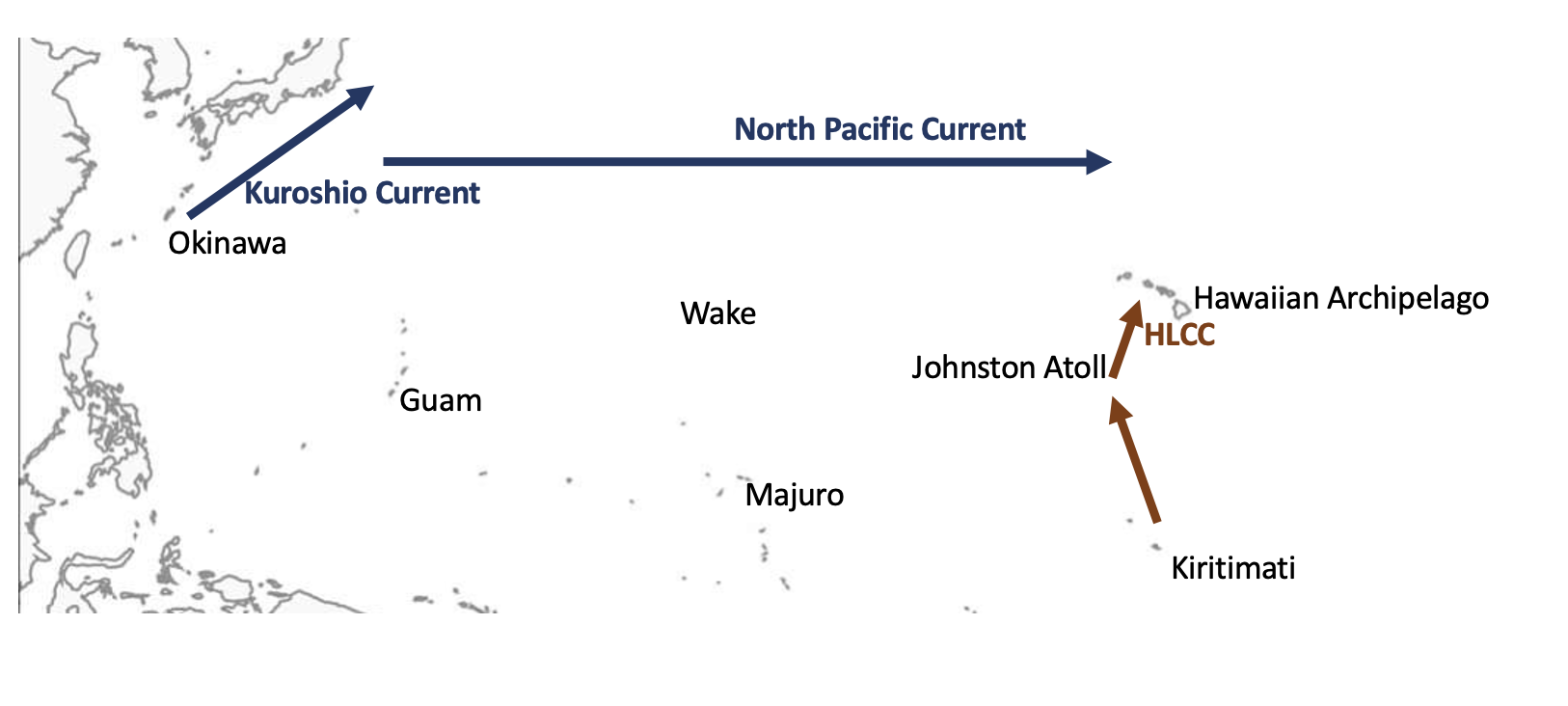
The Hawaiian Archipelago is isolated not just in terms of distance to the nearest archipelago but also because there is no direct current that can quickly transport larvae from surrounding archipelagos. Despite this isolation, species with planktonic larvae have the potential for large dispersal. There are two main hypotheses for the origin of species in Hawaiian Archipelago. One proposed source is the Central Pacific (Line islands through Johnston Atoll) from which larvae are transported via the Hawaiian Lee countercurrent (HLCC). The other proposed source of larvae to the Hawaiian archipelago is southern Japan from which larvae are transported via the Kuroshio Current and then the north Pacific Current.
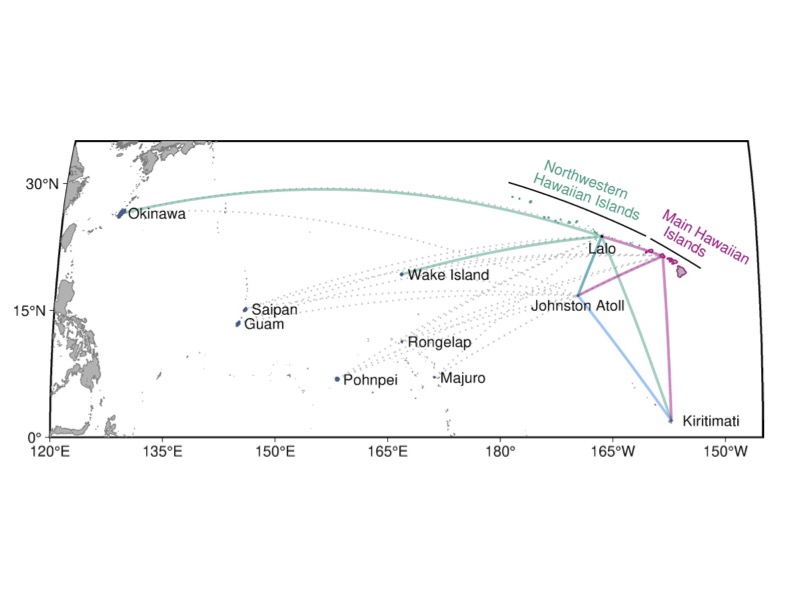
To investigate current connectivity of marine species, I will model larval dispersal potential with varying realistic larval behavior. We have finished a sensitivity analysis for this model and published as a NOAA technical report. We are currently expanding the simulation over 10 years and 6 species with varying life histories to identify the range of dispersal potential between archipelagos. From the results I will be able to identify which life histories are likely to result in connectivity among archipelagoes. I will then compare the connections found in the model to those found using eDNA methods here. Biophysical larval dispersal models are a powerful tool that allows for an estimate of contemporary dispersal potential. On the other hand, genetic data can typically only detect older breaks (greater than ~50,000 years) in genetic connectivity. If barriers to dispersal have appeared within the last ~50,000 years, there is often not enough time for genetic drift alone to cause significant genetic differentiation between sites.